Aminoácidos
1. Características estructurales comunes
Tienen
en común la existencia de un átomo de carbono (carbono α)
al que se une un grupo carboxilo, un grupo amino y un átomo de hidrógeno. La
cuarta valencia del carbono está unida a una cadena lateral diferente en cada aminoácido. La prolina es una
excepción al tener una estructura cíclica.
Excepto
la Gly, el carbono α tiene cuatro sustituyentes distintos, por lo que es un
centro quiral. Por lo tanto existen enantiómeros
que pueden ser D y L. Los aminoácidos que forman las proteínas sólo son L. Las
formas D se pueden encontrar excepcionalmente en parte de la pared bacteriana.
2. Los aminoácidos se clasifican por las características de su cadena lateral
Las
cadenas laterales pueden ser hidrófobas (apolar), polar sin carga o presentar
carga a determinados pH.


3. Los aminoácidos presentan carácter ácido o básico
Los
aminoácidos presentan un grupo amino de carácter básico (aceptor de protones) y
un grupo carboxilo con carácter ácido (dador de protones). A pH fisiológico se
encuentran en forma de ión dipolar. Este tipo de sustancias que pueden
comportarse como ácidos y bases se conocen como sustancias anfóteras. El punto en el cual la carga neta de la molécula sea
cero se conoce como punto isoeléctrico.
4. La carga eléctrica de los aminoácidos varia con el pH
Tomando
como ejemplo la Gly, valores por debajo del pH del pK del grupo ácido (pK1=2,34)
domina la forma totalmente protonada con carga neta positiva. Por encima de
este valor, empieza a aumentar el número de cargas que se encentran en ión
dipolar hasta llegar al punto isoeléctrico. Por encima de este valor, el grupo
NH3+ va perdiendo protones, hasta que predominará la forma con carga
negativa (por encima de pK2=9.6).
Los
aminoácidos con cadenas laterales con grupos ionizables tienen curvas de titulación
con tres valores de pKa. Por tanto, los puntos isoeléctricos se
calcularán como la media aritmética de los dos pKa de los grupos ácidos
o básicos.
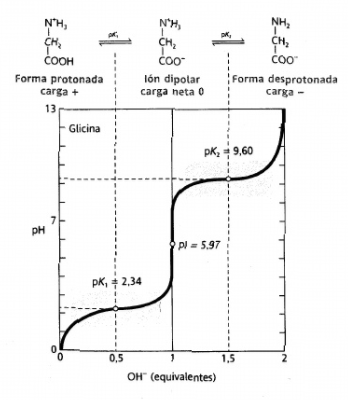
5. Existen regiones pH en que los aminoácidos presentan poder tamponante
En
las zonas próximas a los valores de pKa de ambos grupos (COOH y NH3+)
se necesita más cantidad de base (NaOH) para que se produzca un aumento de pH
de la solución (ver anterior imagen). Este efecto se conoce como efecto tampón.
Solo la His puede actuar a pH cercano al fisiológico. En la hemoglobina, los protones liberados
durante el metabolismo se unen a un residuo de His evitando la disminución de pH,
y cambiando además, su afinidad por el oxígeno.
6. Comportamiento ácido-base de las cadenas laterales
Ciertos
aminoácidos (Lys, Arg, His, Asp, Glu, Cys y Tyr) poseen grupos funcionales con
carácter ácido o básico. El estado ionizado va a depender del valor de su pKa.
A pH fisiológico, todos se encontrarán en su forma ácida excepto la His.
La
importancia biológica de estos grupos radica en su posibilidad de participar en
las reacciones enzimáticas mediantes dos procesos. En primer lugar, estos
aminoácidos pueden participar como aceptores y dadores de protones. En segundo
lugar, los ácidos pueden actuar como reactivos nucleofílicos y las bases como
reactivos electrofílicos en reacciones de ruptura y formación de enlaces.
7. Las propiedades de las cadenas laterales determinan las interacciones no covalentes
7.1
Hidrofobicidad
Muchos
aminoácidos presentan una cadena lateral totalmente apolar, lo que provoca
repulsión, plegándose así su estructura tridimensional formando un núcleo
hidrofóbico.

7.2
Fuerzas de Van der Waals
En
las cadenas laterales de los aminoácidos apolares se pueden crear dipolos
instantáneos. Así se establecen fuerzas de atracción intermoleculares muy
débiles.

7.3
Formación de puentes de hidrógeno
Algunas
cadenas puede actuar como donadoras o receptoras de hidrógeno.

7.4
Interacciones electrostáticas
Las
cadenas laterales pueden estar ionizadas a determinados pH. Dos cadenas laterales
de signos opuestos se atraerán, creando interacciones no covalentes (enlaces salinos).

8. Unión de elementos metálicos
La
unión de cationes divalentes (Mg2+, Ca2+, Zn2+)
y algunos metales de transición (Fr, Cu, Ni…) es bastante frecuente en muchas
enzimas u otras proteínas como la hemoglobina. Estos metales se unen formando
entidades de coordinación, que están compuestas por un átomo central (metal)
que está unido a átomos denominados ligandos. En el grupo hemo, el Fe2+ establece cuatro enlaces con los átomos
del N del anillo de porfrina. El quinto enlace se forma con el N del grupo imidazol
de una histidina (His proximal). El sexto sólo se forma en presencia de O2.

Los
aminoácidos con cadenas laterales ionizables pueden establecer interacciones
con elementos metálicos mediante interacciones no covalentes iónicas.
9. Las cadenas laterales pueden establecer enlaces covalentes
9.1
Formación de puentes disulfuro
Dos
residuos de Cys pueden unirse covalentemente por enlace disulfuro mediante un
proceso de oxidación. Pueden ser en la misma proteína (intracatenarios) o entre dos cadenas diferentes (intercatenarios).
Al requerir
un ambiente oxidante no son comunes en el citosol (ambiente reductor). Sin
embargo, si lo son en las proteínas extracelulares que son secretadas por la
célula. La formación de estos enlaces se produce en el lumen del retículo
endoplasmático.
Este
tipo de enlace mantiene la estructura tridimensional de las proteínas. Es muy
común en las inmunoglobulinas.

9.2
Fosforilación
Las
cadenas laterales de Ser, Thr y Tyr pueden sufrir fosforilación por la unión de
un grupo fosfato inorgánico a su grupo hidroxilo. Este proceso está catalizado
por las quinasas y puede ser
reversible por las fosfatasas.
La
adición de un grupo fosfato con carga negativa, hace que se produzcan cambios
conformacionales, lo que afecta directamente en la actividad. Puede inactivarla
o activarla.
9.3
glucosilación
Los
glúcidos se pueden unir de forma covalente a las proteínas mediante la
formación de un enlace O-glucosídico o N-glucosídico.

10. La estereoquímica de las cadenas laterales es fundamental para comprender la estructura tridimensional de las proteínas
Las
características químicas, geometría y distribución espacial de los enlaces de
átomos que forman las cadenas laterales, van a tener importancia para determinar
el plegamiento de las proteínas y sus interacciones con otras moléculas.
11. Modificaciones químicas de los aminoácidos en las proteínas
Existen
aminoácidos que sufren modificaciones después de estar incorporados en la
cadena polipeptidica. Ejemplos son la 4-hidroxiprolina
y la 5-hidroxilisina, que se
encuentran en la molécula de colágeno (adicción de grupo hidroxilo). Existen
cambios más complejos como la formación de desmosina
en la elastina.


Algunos
aminoácidos de las proteínas que forman complejos con los ácidos nucleicos (histonas) pueden tener grupos metilo,
acetilo o fosfato específicos.
Otro
ejemplo es el ácido ϒ-carboxiglutámico,
que se encuentra la protrombina, una de las proteínas que intervienen en la
coagulación de la sangre.
12. Aminoácidos no presentes en las proteínas
Es
frecuente que algunos derivados de aminoácidos funcionen como mensajeros
químicos. Por ejemplo GABA (ácido ϒ-aminobutírico), que se produce por la
descarboxilación del glutamato. Es un neurotransmisor con función inhibitoria,
y es un blanco frecuente en fármacos que reducen la ansiedad como los
barbitúricos (en desuso).Otro ejemplo es la dopamina, derivado de la Tyr y un neurotransmisor relacionado con
la enfermedad de Parkinson (descenso de dopamina).
No hay comentarios:
Publicar un comentario