La histología, también llamada
anatomía microscópica, es el estudio de las estructuras microscópicas de los
tejidos y órganos del cuerpo.
Para ello, en la actualidad, se
emplean más comúnmente el microscopio óptico, el de fuerza atómica (MFA) y
además la microscopía virtual (método que permite examinar la muestra en la
pantalla de un ordenador o dispositivo móvil).
2. Preparación del tejido
Tinción con hematoxilina y
eosina con fijación en formalina
Esta técnica es la que se utiliza
con mayor frecuencia.
Primer paso: La fijación. Se realiza para conservar la estructura
de la muestra, para poder llevar a cabo los tratamientos posteriores. La
muestra debe sumergirse en el fijador nada mas extraerse. El fijador
comúnmente usado es la formalina: solución acuosa de formaldehido al 37%, en
combinación con otras sustancias químicas.
Sin embargo, la formalina, no
reacciona con los lípidos, por lo que es un mal fijador de las membranas
celulares. Así pues, para conservar las estructuras de la membrana se emplean
fijadores especiales, con metales pesados que se unan a los fosfolípidos, tales
como permanganato y osmio.
Segundo paso: después de dicha fijación, la muestra se prepara para
su inclusión en parafina. Se lava y se deshidrata mediante disoluciones
alcohólicas de concentración creciente, hasta llegar al 100%. A continuación la
muestra se aclara, con solventes orgánicos, tales como el xileno o el tolueno,
con el objetivo de extraerle el alcohol.
A continuación se realiza la
inclusión de la muestra en parafina fundida. Se realiza con el fin de poder
realizar posteriormente un corte muy delgado de la muestra (de 5 a 15 µm).
Una vez que la parafina se ha secado y endurecido se formará un bloque, en cuyo
interior se encuentra la muestra. Dicho bloque se colocará en una máquina
cortadora, llamada micrótomo, que realizará los cortes ya mencionados. Estos
cortes se colocan sobre un portaobjetos de vidrio, utilizando pineno o resinas
de acrílico como adhesivo.
Tercer paso: tinción de la muestra, con el objetivo de permitir su
posterior examen, puesto que los cortes de parafina son incoloros. Para ello, la
parafina debe extraerse con xileno o tolueno y los tejidos han de
rehidratarse, mediante soluciones de alcohol de concentración decreciente. A continuación el tejido que ha quedado sobre
el portaobjetos se tiñe con hematoxilina en agua. Después, la muestra se vuelve
a deshidratar, mediante las soluciones de alcohol de concentraciones crecientes
y se vuelve a teñir con eosina en alcohol.
Cuarto paso: la muestra se pasa por xileno o tolueno y se le coloca
un medio de montaje no acuoso. A continuación se le coloca un cubreobjetos para
obtener así el preparado final y permanente.
Demostración del efecto de la
hematoxilina y la eosina actuando por separado y juntas:
Ambos colorantes juntos van a
conseguir que la muestra se pueda examinar con mucha mayor precisión, ya que
cada uno por separado tiñe los elementos de la muestra para los que presenta
más afinidad.
Otras tinciones
A pesar de los buenos resultados
obtenidos con la tinción de hematoxilina y eosina, algunos componentes, tales
como la elastina, fibras reticulares, membranas basales y lípidos, no se pueden
observar claramente. Por ello, cuando se desea examinar dichos componentes, es
necesario emplear procedimientos de tinción selectivos. Dichos procedimientos incluyen el uso de
orceína y fucsina-resorcina, para los materiales elásticos, y la impregnación
argéntica para las fibras reticulares y membranas basales.
Detection and localization of
surgically resectable cancers with a multi-analyte blood test
Un reciente estudio describe una nueva prueba de diagnóstico precoz
para el cáncer. A través de ella se podrían detectar ocho tipos comunes de
cáncer, simplemente evaluando los niveles de proteínas circulantes y las
mutaciones en el ADN libre de las células.
La prueba llamada CancerSEEK, fue utilizada en 1005 pacientes con
cánceres no metastásicos en: ovario, hígado, estómago, páncreas, esófago, colon,
pulmón o mama.
Los resultados estadísticos son bastante variables. La media de
detección fue del 70% en los ocho tipos de cáncer. La sensibilidad fluctuó del
69% al 98% en los cánceres en los que no existen pruebas de detección precoz disponibles para individuos en riesgo (ovario, hígado, estómago, páncreas y
esófago).
En cuanto a la especificidad del CancerSEEK fue mayor del 99%. De
los 812 controles sanos se obtuvieron únicamente 7 falsos positivos.
Bibliografía:
Cogen JD, Li L, Wang Y, Thoburn C, Afsari B, Danilova L, et al. Detection and localization of surgically resectable
cancers with a multi-analyte blood test. Sciente. 2018;359.
2.1 Los volúmenes pulmonares
cambian durante la ventilación
Para evaluar la función pulmonar
de una persona se realizan pruebas de función pulmonar con un espirómetro. Los volúmenes pulmonares pueden dividirse en 4:
Volumen
corriente o tídal (Vc): Aire que se mueve durante una respiración normal no forzado (inspirado y espirado).
Suele ser de 500 mL.
Volumen
de reserva inspiratoria (VRI): Es el aire extra, además del volumen corriente, que podemos meter forzadamente en los
pulmones. Suele ser de unos 3000 mL.
Volumen
de reserva espiratoria (VRE): Es la cantidad de aire que se puede exhalar forzadamente, además del volumen corriente. Ronda los 1100 mL.
Volumen
residual (VR): Es el volumen de aire que queda dentro de los pulmones
después de una espiración máxima. No se puede expulsar y su función es
mantener los pulmones estirados y pegados a las costillas. Ronda los 1200
mL.
Las capacidades pulmonares son la suma de dos o más volúmenes
pulmonares. Tenemos:
Capacidad
vital (Vc + VRI + VRE): Es la cantidad máxima de aire que puede eliminarse tras una espiración máxima. Es igual a la suma de los volúmenes de reserva
inspiratoria, espiratoria y corriente.
Capacidad
pulmonar total (Vc + VRI + VRE + VR): Es la cantidad total de aire en los pulmones tras una inspiración máxima. Es igual a la suma de capacidad vital y volumen
residual.
Capacidad
inspiratoria (Vc + VRI): Es la cantidad máxima de aire que puede inspirarse tras una espiración normal. Es igual a la suma del volumen corriente y volumen de reserva
inspiratoria.
Capacidad
funcional residual (VRE + VR): Es la cantidad de aire que permanece en los pulmones tras una espiración normal. Es igual a la suma del volumen de reserva espiratoria y volumen
residual.
2.2 La inspiración se produce
cuando la presión alveolar disminuye
Para que el aire fluya hacia los
pulmones, la presión dentro de ellos debe ser más baja que la atmosférica.
Cuando el diafragma se contrae, el abdomen desciende (60-75% del cambio del
volumen). El movimiento de la caja torácica mediado por los intercostales externos y
escalenos provoca el 25-40% del cambio de volumen. A medida que el volumen
torácico aumenta, las presiones disminuyen y el aire fluye hacia el interior.
Si los escalenos de un individuo
están paralizados, la inspiración se logra con el diafragma, en mayor parte. En
el caso de los intercostales externos, a penas participan en las respiraciones
en reposo, pero ganan importancia a medida que aumenta la actividad
respiratoria.
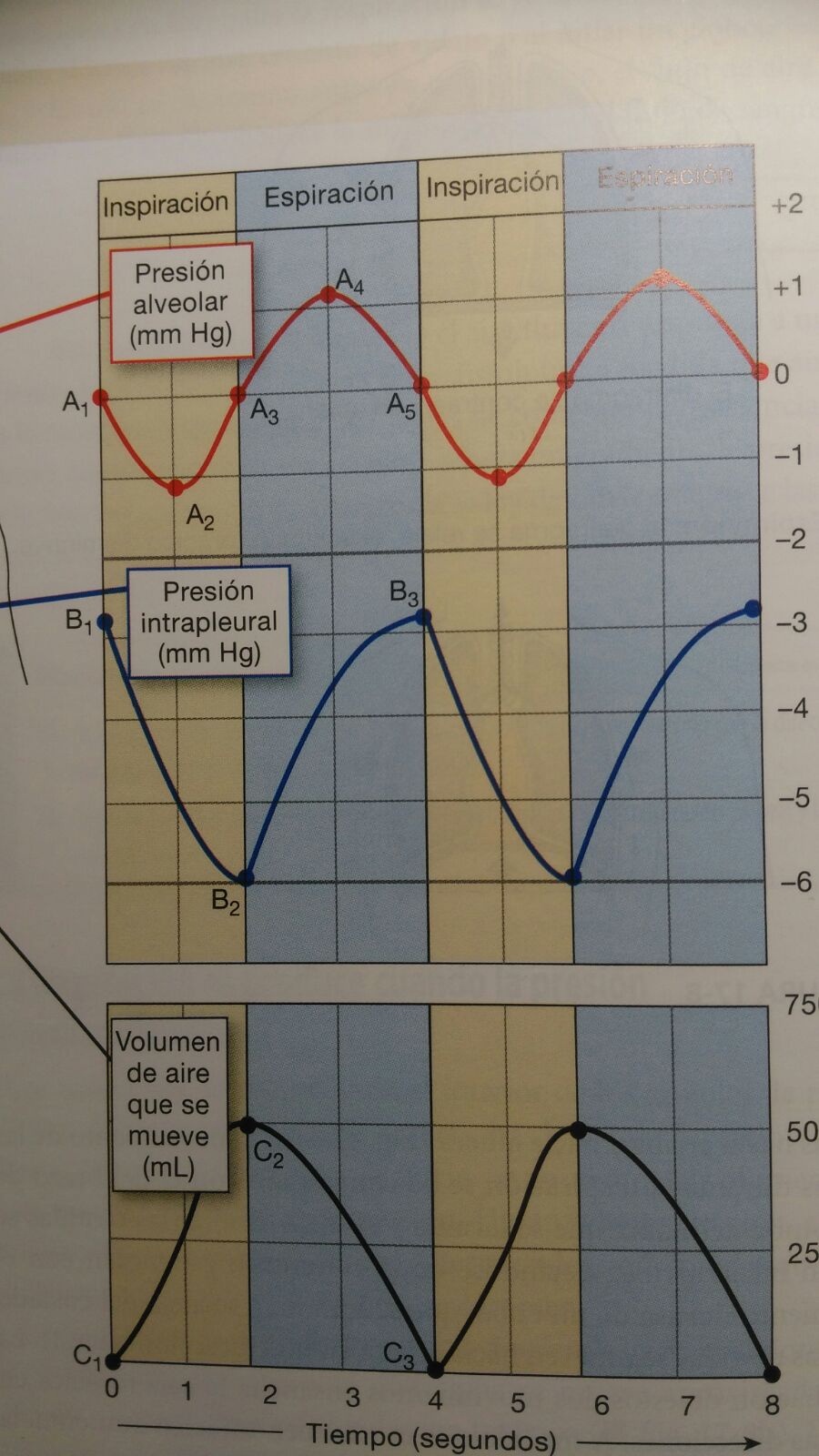
Si decimos que la presión
atmosférica es 0 mmHg, para mayor facilidad del estudio, tenemos varios tiempos
de inspiración con determinadas presiones:
Tiempo
0s: Breve pausa entre respiraciones donde la presión alveolar es igual a
la atmosférica (0 mmHg en A1).
Tiempo
0 a 2s: Se produce la inspiración con la contracción de los músculos
respiratorios. La presión disminuye 1 mmHg (A2) y el aire fluye
hacia dentro. Como el volumen torácico cambia más rápido de lo que puede
fluir el aire, la presión alveolar alcanza su valor más bajo
aproximadamente a la mitad de la inspiración.
Al final de la inspiración, la
presión alveolar es igual a la atmosférica (A3) y el volumen
pulmonar alcanza el máximo del ciclo ventilatorio (C2).
2.3 La espiración se produce
cuando la presión alveolar aumenta
Al final de la espiración, los
impulsos de las motoneuronas somáticas hacia los músculos inspiratorios
cesan. La recuperación elástica lleva al
diafragma y la caja torácica a sus posiciones relajadas originales.
Tiempo
2-4s: A medida que el volumen pulmonar disminuye, la presión alveolar
aumenta 1 mmHg (A4) por encima de la atmosférica lo que provoca
que el aire salga.
Tiempo
4s: Al final de la espiración, el movimiento de aire cesa cuando la
presión alveolar se iguala con la presión atmosférica (A5) . El
volumen pulmonar alcanza su valor mínimo del ciclo ventilatorio (C3).
En la respiración activa (más de 30 respiraciones por minuto) se utilizan
los músculos intercostales internos y abdominales (músculos espiratorios). Los
músculos abdominales se contraen tirando de la caja torácica hacia dentro y
disminuye el volumen abdominal, acciones que desplazan los intestinos y el
hígado hacia arriba. Las vísceras empujan el diafragma hacia arriba, lo que
disminuye aún más el volumen pulmonar.
Cualquier enfermedad
neuromuscular que debilite los músculos esqueléticos o dañe sus motoneuronas
puede afectar de manera adversa a la ventilación. Por ejemplo, la miastenia grave cursa con destrucción de receptores de acetilcolina
de las placas terminales motoras de los músculos esqueléticos. La poliomielitis es una enfermedad viral
que paraliza los músculos esqueléticos.
2.4 La presión intrapleural
cambia durante la ventilación
La superficie de los pulmones
está recubierta por la pleura visceral
y la cavidad torácica por la pleura
parietal. Las dos membranas se mantienen unidas por la presencia del
líquido pleural, de forma que los pulmones elásticos son forzados a estirarse
para ocupar el mayor volumen. Al mismo tiempo, la recuperación elástica de los
pulmones genera una fuerza hacia dentro (inversa). La combinación de la fuerza
hacia fuera de la caja torácica y la recuperación hacia dentro de los pulmones
elásticos genera una presión intrapleural subatmosférica de -3 mmHg.
Una entrada de aire a la cavidad
pleural, rompe el enlace líquido que mantiene el pulmón unido a la pared
torácica. El pulmón por tanto se colapsa, eso se conoce como neumotórax.
A modo de curiosidad, del 3 al 5%
del gasto de energía del cuerpo se utiliza para la respiración en reposo.
2.5 La distensibilidad y la
elasticidad de los pulmones pueden cambiar en los estados patológicos
La capacidad de estirarse de los
pulmones se conoce como distensibilidad,
mientras que lo opuesto, la capacidad para resistir a la deformación, se conoce
como elasticidad.
La disminución de la
distensibilidad provoca que deba ejercerse más trabajo para estirar un pulmón
rígido. Esto ocurre en las enfermedades
pulmonares restrictivas. Otras dos causas comunes de disminución de la
distensibilidad son el tejido cicatricial no elástico formado en las enfermedades fibróticas pulmonares y
la producción alveolar inadecuada de surfactante.
La fibrosis pulmonar se caracteriza por el desarrollo de tejido
cicatricial rígido y fibroso que restringe el inflado del pulmón. Puede ser de
forma idiopática o causada por la inhalación crónica de partículas finas como
asbestos o sílice. En este caso, los macrófagos no pueden digerirlas (porque no
son moléculas orgánicas), por lo que se acumulan, y se desencadena un proceso
inflamatorio. Los macrófagos secretan factores de crecimiento que estimulan los
fibroblastos del tejido conectivo pulmonar, para producir colágeno no elástico (no
se puede revertir el proceso).
2.6 El surfactante disminuye
el trabajo de la respiración
El líquido que recubre los
alvéolos genera tensión superficial. En los pulmones, el surfactante disminuye
esta tensión y, por lo tanto, reduce la resistencia del pulmón al estiramiento.
El surfactante está más
concentrado en los alvéolos más pequeños, debido a que estos tienen más tensión
superficial (Ley de LaPlace). Este compuesto está formado por una mezcla de
proteínas y fosfolípicos, como dipalmitoilfosfatidilcolina, que es secretada
por las células alveolares del tipo II.
Los bebés prematuros a veces no
tienen concentraciones adecuadas de surfactante y desarrollan el síndrome de dificultad respiratoria del
recién nacido. Estos bebés utilizan mucha energía para expandir sus
pulmones y, si no se trata, puede ser fatal. Actualmente, se administra
surfactante en aerosol y se utiliza ventilación artificial (ventilación de
presión positiva). A través del líquido amniótico se podrá diagnosticar un
déficit en la producción de surfactante en el feto y, de ser así y, si las
condiciones lo permiten, se podrá retrasar el nacimiento.
2.7 El diámetro de las vías
aéreas determina su resistencia
Casi el 90% de la resistencia de
las vías aéreas puede atribuirse a la tráquea y bronquios. Como estas
estructuras están sostenidas por cartílago y hueso, sus diámetros no cambian.
La acumulación de moco aumentará marcadamente la resistencia.
Los bronquiolos no contribuyen,
normalmente, a la resistencia. Sin embargo, una disminución importante o
generalizada del diámetro, puede convertirse en una fuente importante de
resistencia al flujo. Esto se conoce como broncoconstricción.
Los bronquiolos están sujetos al
control reflejo del sistema nervioso y de las hormonas. Sin embargo, la mayoría
de los cambios del diámetro se deben a las sustancia paracrinas. El CO2
es el que más influye, ya que un aumento de este, relaja el músculo liso
bronquial provocando broncodilatación.
Otra sustancia paracrina, la histamina,
es liberada por los mastocitos en respuesta al daño tisular o a las reacciones
alérgica. Una liberación puede provocar broncoconstricción.
El principal control neurológico
(acetilcolina en receptores muscarínicos) proviene de las neuronas
parasimpáticas que causan broncoconstricción, un reflejo diseñado para proteger
de sustancias irritantes inhaladas el tracto respiratorio inferior. No hay
inervación simpática significativa, sin embargo, en el músculo liso de los
bronquiolos hay receptores β2
que responden a la adrenalina. La estimulación de estos receptores relaja el
músculo liso y provoca broncodilatación (útil en asma o reacciones alérgicas).
2.8 La frecuencia y la
profundidad de la respiración determinan su eficacia
Es importante conocer varias
fórmulas:
Ventilación
pulmonar total = F. Respiratoria x volumen corriente (Unos 6L/minuto)
Ventilación
alveolar = F. Respiratoria x (volumen corriente – espacio muerto) (unos
4200mL/minuto)
Hay que saber que no todo el aire
que entra en el aparato respiratorio llega hasta los alvéolos, porque parte del
aire queda en las vías aéreas de conducción (espacio muerto anatómico), unos 150mL. Por eso, el indicador más
fiable de la eficacia de la ventilación es la ventilación alveolar.
2.9 La composición de los
gases en los alvéolos varía poco durante la respiración normal
La hiperventilación aumenta la PO2
y disminuye la PCO2 en los alvéolos. Sucede lo contrario durante la
hipoventilación. Sin embargo, durante la respiración normal, estas presiones
cambian poco como se cabría esperar. La PO2 suele ser constante en
100 mmHg y la PCO2 suele mantenerse en 40 mmHg. ¿Por qué?
La
cantidad de oxígeno que entra en los alvéolos es casi igual a la cantidad
que entra en la sangre (uno entra y se sustituye por el nuevo).
La
cantidad de aire fresco que entra en los pulmones en cada ciclo
ventilatorio es apenas un poco mayor que el 10% del volumen pulmonar total
al final de la inspiración.
2.10 La ventilación y el flujo
de sangre se corresponden
En una persona en reposo, algunos
lechos capilares del vértice (parte superior) del pulmón están cerrados debido
a la baja presión hidrostática. Durante el ejercicio, se reclutan para asegurar
que el organismo está oxigenado.
A nivel local, el organismo
intenta equiparar el flujo de aire y sangre variando el diámetro de las arteriolas
y bronquiolos. El diámetro bronquiolar está mediado por la concentración de CO2,
y el diámetro de las arteriolas pulmonares está regulado principalmente por el
contenido de oxígeno de líquido intersticial que rodea la arteriola.
Un ejemplo práctico: Si la
ventilación en un alvéolo disminuye, la PO2 de esa zona también. Se produce vasoconstricción para dirigir la sangre hacia otros alvéolos mejores
ventilados.
Los
mecanismos de regulación no son eficaces en sucesos como bloqueo de arteria
pulmonar o bloqueo de vías aéreas superiores, porque este mecanismo no podría
mejorar la situación.
2. 11 La auscultación y la
espirometría evalúan la función pulmonar
La auscultación de los sonidos
respiratorios es una técnica diagnóstica importante en la medicina pulmonar.
Cuando el flujo de aire disminuye, los sonidos están ausentes o se aprecian poco.
La inflamación de las membranas pleurares produce un sonido crepitante o áspero, conocido como frote pleural.
En las enfermedades obstructivas crónicas se escucha un silbido del aire a
través de las vías aéreas. En las EPOC se incluye el asma, la apnea obstructiva
del sueño, el enfisema y la bronquitis crónica. En la apnea obstructiva del sueño se produce obstrucción de las vías
aéreas superiores por relajación anormal de los músculos de la faringe y la
lengua. El asma es una enfermedad
inflamatoria que produce broncoconstricción y edema de las vías aéreas. La broncoconstricción,
en este caso, es causada por acetilcolina, histamina, la sustancia P (un
neuropéptido) y los leucotrienos secretados por los mastocitos, macrófagos y eosinófilos.
Esta patología se trata con β2-adrenérgicos,
antiinflamatorios y antagonistas de leucotrienos.
Comenzamos hablando de las principales funciones del aparato
respiratorio. Son las siguientes:
El intercambio de gases entre la atmósfera y la
sangre.
La regulación homeostática del pH del organismo.
La protección frente a los patógenos inhalados y las
sustancias irritantes.
La vocalización.
1. El aparato
respiratorio
La respiración
celular es la reacción intracelular del oxígeno con moléculas orgánicas, para producir dióxido de carbono, agua y energía en forma de ATP. La respiración externa es el movimiento de
gases entre el medio ambiente y las células del cuerpo. Se divide en 4
procesos:
El intercambio de aire entre la atmósfera y los
pulmones.
El intercambio de O2 y CO2
entre los pulmones y la sangre.
El transporte de estos gases a través de la sangre.
El intercambio de gases entre la célula y la
sangre.
El aparato respiratorio está formado por las vías aéreas, los
alvéolos y sus capilares asociados, y los huesos y músculos que asisten la
ventilación.
1.1 Los huesos y los músculos del tórax rodean los
pulmones
Al conjunto se le denomina caja torácica. Las costillas y la
columna forman los costados y la parte superior de la caja. Una lámina de músculo
esquelético con forma de cúpula, el diafragma,
forma la parte inferior.
Los músculos intercostales
conectan los 12 pares de costillas. Los esternocleidomastoideos
y los escalenos van desde la cabeza y
cuello hasta el esternón y las dos primeras costillas.
El tórax es un contenedor sellado formado por el saco
pericárdico, que contiene el corazón, y los sacos pleurales que rodean a cada uno de los pulmones.
1.2 Los sacos pleurales contienen los pulmones
Las vías aéreas semirrígidas (bronquios) conectan los pulmones
con la vía aérea principal. Cada pulmón está rodeado por un saco pleural, de
pared doble, cuyas membranas recubren la parte interior del tórax y la
superficie externa de los pulmones. Cada pleura
contiene varias capas de tejido conectivo y numerosos capilares. Las capas opuestas de la pleura se mantienen
juntas por el líquido pleural.
Este líquido cumple varios propósitos:
Crea una superficie húmeda y resbaladiza para
que las membranas opuestas puedan deslizarse.
Mantiene unidos los pulmones a la pared torácica, debido a la fuerza de cohesión del agua.
1.3 Las vías aéreas conectan los pulmones con el medio
externo
El aire entra por la boca y la nariz, y pasa a la faringe; después a la laringe y a la tráquea. En la laringe se encuentran las cuerdas
vocales.
La tráquea es un tubo semiflexible y abierto, por la presencia
de 15-20 anillos de cartílago con forma de C. Se ramifica en bronquios primarios, y estos repetidamente
se dividen. Posteriormente se dividen en bronquíolos,
que se siguen ramificando, hasta formar una transición entre las vías aéreas y
el epitelio de intercambio.
1.4 Las vías aéreas calientan, humedecen y filtran el
aire inspirado
El acondicionamiento tiene tres fases:
Calentar el aire a la temperatura del cuerpo,
para que los alvéolos no se dañen y la temperatura corporal no cambie.
Agregar vapor de agua hasta humedecer al 100%.
Filtrar todos los elementos extraños.
Las vías aéreas (tranquea y bronquios) están recubiertas de
epitelio ciliado, cuyas células están bañadas por una capa de solución salina
acuosa. Dicha capa es elaborada por las células epiteliales, cuando el Cl-
es secretado hacia la luz por los canales aniónicos apicales. Conscuentemente, esto arrastra el
Na+ a través de una vía paracelular. Se creará así un gradiente
osmótico, y el agua sigue a los iones hacia la luz de las vías aéreas.
La capa mucosa es secretada por las células caliciformes del
epitelio. Los cilios arrastran el moco hacia la parte superior, lo que se
denomina transporte mucociliar. El
moco contiene inmunoglobulinas que ayudan en la defensa.
La secreción de una capa de solución salina es esencial para
el transporte mucociliar. Una secreción inadecuada de iones provocará que no se
forme la solución salina, el moco se espesará y no podrá ser eliminado. Así pues, las bacterias podrán colonizar las vías aéreas y provocar infecciones pulmonares
recurrentes.
1.5 Los alvéolos son el sitio de intercambio de gases
Cada pequeño alvéolo está formado por una sola capa de
epitelio. Las células alveolares de tipo
II sintetizan el surfactante,
que ayuda al pulmón a expandirse. Estas células también ayudan a minimizar la
cantidad de líquido presente en los alvéolos. Las células alveolares de tipo I (95%), son las que participan en la
difusión de gases.
Las delgadas paredes de los alvéolos no contienen músculo, ya que si estuviera presente bloquearía el intercambio de gases. Por lo tanto, la contracción se debe
a la propia capacidad elástica que está provocada por las fibras de elastina y
de colágeno.
1.6 La circulación pulmonar tiene alto flujo y baja
presión
Comienza en el tronco de la arteria pulmonar, que recibe
sangre con poco oxígeno del ventrículo derecho. La sangre regresa al corazón
por las venas pulmonares.
La velocidad de flujo a través de los pulmones es mucho más
alta que en otros tejidos, pero esto no provoca aumento de la presión arterial.
El valor promedio de presión es de 25/8 mmHg, en comparación con los 120/80 mmHg sistémicos. Esto es consecuencia de que el ventrículo derecho no tiene que bombear con fuerza, porque las resistencias
pulmonares son bajas, debido a la corta longitud de los vasos sanguíneos, su
distensibilidad y el gran área de sección transversal total de las arteriolas
pulmonares.
En condiciones normales la presión hidrostática es baja, por lo que la cantidad de líquido extravasado es escaso. Así pues, en estas condiciones, el sistema linfático es capaz de eliminar ese líquido.
La superficie celular y la matriz
extracelular están compuestas por moléculas ricas en glúcidos. Al hablar de glucoconjugados, dichas moléculas se unen a proteínas o lípidos (glucoconjugado), mediante enlace covalente. El papel de los glúcidos es transportar
información: actúan en el reconocimiento y adhesión celular, migración celular,
respuesta inmunitaria… Los glucolípidos
son lípidos de membrana (esfingolípidos) unidos a oligosacáridos, las glucoproteínas son proteínas unidas a
oligosacáridos y los proteoglucanos son proteínas unidas a polisacáridos glucosaminoglucanos (GAG).
El hecho de que los
glucoconjugados estén siempre orientados hacia el exterior celular viene
determinado por su síntesis, ya que los oligosacáridos se adicionan durante el
transporte, desde su lugar de síntesis en el retículo endoplasmático y su paso
por el aparato de Golgi.
Proteoglucano formado por una proteína transmembrana
Los diferentes GAG generan una
gran variedad de proteoglucanos
En los mamíferos se pueden
encontrar hasta 30 tipos diferentes de proteoglucanos. La unidad básica es una
proteína núcleo a la que se unen por enlace covalente 1 o más GAG.
Existen varias familias de
proteínas núcleo:
Sindecán: Proteínas transmembrana
que tienen unido el sulfato de condroitina y el sulfato de heparán.
Glupicanos: También están anclados
a la membrana, pero a través de un lípido de membrana.
Algunas veces se pueden formar
grandes agregados de proteoglucanos. Un ejemplo es el agrecán, que proporciona consistencia, resistencia y tensión a la matriz del
tejido conjuntivo de los cartílagos. Este compuesto está formado por una molécula de ácido
hialurónico, a la que se unen de forma no covalente 100 proteínas enlace. A su
vez, se unirán a más agrecán que
estará unidos a diferentes GAG.
Las glucoproteínas contienen
oligosacáridos ricos en información
La adición de los oligosacáridos
se realiza durante el proceso de síntesis de proteínas del retículo
endoplásmico.
Las glucoproteínas siempre van a
tener unidos covalentemente oligosacáridos a un determinado aminoácido de la
proteína:
Si los
glúcidos se unen al aminoácido asparagina (Asn) la unión será de tipo N-glucosídico, ya
que este aminoácido aporta el grupo NH2 al enlace con el
carbono anomérico del glúcido.
Si se
unen a la serina (Ser) o a la treonina (Thr) la unión será O-glucosídica, porque el aminoácido
aporta el grupo –OH.
La proporción glúcídica de las
glucoproteínas es mucho menor que la de los proteoglucanos, sin embargo, son compuestos
mucho más complejos. Esto se debe a que los oligosacáridos pueden estar
compuestos por monosacáridos diferentes y a que también pueden presentar muchas
ramificaciones. Además, pueden ser modificados por grupos sulfato lo que aporta
información a la molécula.