3. El ADN
3.1 Las moléculas de ADN tienen diferente composición de bases
La
composición de las bases en el ADN está sujeta a las reglas de Chargaff:
- La composición de las bases del AND varía de un sujeto a otro.
- El ADN dentro de un sujeto es igual en todos los tejidos.
- La composición del ADN no varía con la edad, ni con su estado nutricional, ni con las variaciones ambientales.
- En todas las especies la cantidad de adenina es igual a la de timina (A=T), y también (G=C). Por tanto A+G=T+C, es decir, la suma de los residuos de purinas es igual a la suma de residuos de pirimidinas.
La
estructura del ADN fue descubierta por James Watson y Francis Crick en 1953. Se
trata de dos cadenas de polinucleótidos enrolladas alrededor de un eje común
formando una doble hélice. Las dos
cadenas son antiparalelas con
enrollamiento dextrógiro y presentan
dos surcos, el mayor y el menor. Hay que tener en cuenta que las
bases están orientadas hacia el centro de la hélice, y las pentosas y los
fosfatos hacia el exterior. Cada base está unida por puentes de hidrógeno (apareamiento de bases complementarias)
a una hebra opuesta (par de bases).
La
molécula de ADN se mantiene unida por los enlaces de hidrógeno. Entre G y C se
forman tres enlaces, mientras que entre A y T sólo dos. Un ADN que tenga más G
y C será más difícil de separar.
3.2 El ADN puede adoptar distintas formas tridimensionales
La
estructura “típica” que se estudia normalmente (la descubierta por Watson y Crick)
es denominada forma B. Es la forma
más estable del ADN.
La forma A predomina en disoluciones
pobres en agua. Se trata de una hélice dextrógira pero más gruesa y con 11
pares de bases por vuelta (10,5 en forma B). El plano de las pares de bases
tiene una inclinación de 20º con respecto al eje de la hélice. El surco mayor
es más profundo y el surco menor es más superficial.
La forma Z es una hélice levógira que contiene
12 pares de bases por vuelta. Es una estructura más delgada y alargada.

4. Estructura y función del ARN
El
ARN se produce al transcribir ADN, es decir, el ARN es complementario.
Posteriormente el ARN es traducido a
la secuencia correspondiente de aminoácidos que formarán una proteína. Hay
varios tipos de ARN:
- ARN ribosómino (ARNr): Es el componente principal de los ribosomas y tienen un papel catalítico y estructural en la síntesis de proteínas. Los ribosomas están formados por dos subunidades (mayor y menor) que, para analizarlas, se separan por ultracentrifugación. El movimiento de una partícula en esta técnica se caracteriza por un coeficiente de sedimentación que se expresa en unidades Sverdberg (S). Los ribosomas eucarióticos tienen un coeficiente de sedimentación de 80S, y la subunidad grande y pequeña 60S y 40S respectivamente.
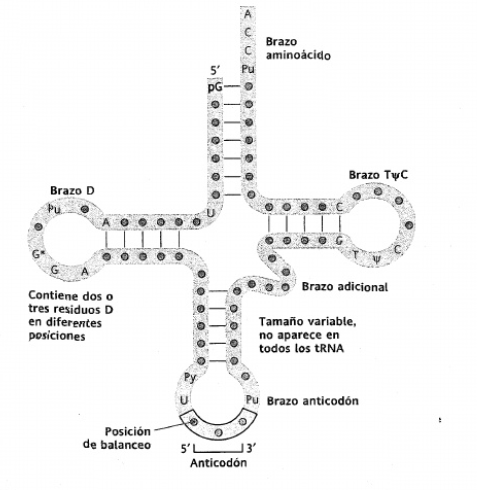
- ARN de transferencia (ARNt): Es muy pequeño y consta de solo una cadena. Su función es transportar los aminoácidos en forma activada hacia el ribosoma. En el ARNt se forman puentes de hidrógeno entre bases igual que en el ADN, pero la timina se sustituye por el uracilo. En su estructura se puede apreciar un brazo del anticodón que contiene el anticodón, un brazo D que contiene dihidrouridina (D), y el brazoT ΨC (brazo T) que contiene ribotimina y pseudouridina.
- ARN mensajero (ARNm): Es el menos abundante y su función es ser el molde para la síntesis de proteínas. El ARNm es complementario al mensaje genético contenido en un segmento específico del ADN. En eucariotas inicialmente está formado por ARN nuclear heterogéneo con largos intrones que al ser eliminados dan ARNm.
- ARN pequeño nuclear (ARNpn): Se encuentra en el núcleo de las células eucariotas y su función es contribuir al procesamiento del ARNm que se transcribe del ADN para dar una forma madura que se pueda exportar del núcleo. Es necesario porque en eucariotas la transcripción se produce en el núcleo, pero la traducción en el citosol.





